All published articles of this journal are available on ScienceDirect.

Aqueous Humor Dynamics: A Review
Abstract
Glaucoma is a family of optic neuropathies which cause irreversible but potentially preventable vision loss. Vision loss in most forms of glaucoma is related to elevated IOP with subsequent injury to the optic nerve. Secretion of aqueous humor and regulation of its outflow are physiologically important processes for maintaining IOP in the normal range. Thus, understanding the complex mechanisms that regulate aqueous humor circulation is essential for management of glaucoma. The two main structures related to aqueous humor dynamics are the ciliary body and the trabecular meshwork (TM). Three mechanisms are involved in aqueous humor formation: diffusion, ultrafiltration and active secretion. Active secretion is the major contributor to aqueous humor formation. The aqueous humor flow in humans follows a circadian rhythm, being higher in the morning than at night. The aqueous humor leaves the eye by passive flow via two pathways - the trabecular meshwork and the uveoscleral pathway. In humans, 75% of the resistance to aqueous humor outflow is localized within the TM with the juxtacanalicular portion of the TM being the main site of outflow resistance. Glycosaminoglycan deposition in the TM extracellular matrix (ECM) has been suggested to be responsible for increased outflow resistance at this specific site whereas others have suggested deposition of proteins, such as cochlin, obstruct the aqueous humor outflow through the TM. The uveoscleral outflow pathway is relatively independent of the intraocular pressure and the proportion of aqueous humor exiting the eye via the uveoscleral pathway decreases with age.
INTRODUCTION
Glaucoma is a heterogeneous group of eye diseases from the viewpoint of pathogenesis and clinical expression. Glauoma is characterized by optic nerve damage, leading ultimately to irreversible blindness. Glaucoma is estimated to affect approximately 70 million people worldwide [1, 2] and more than 2 million people in the USA. In the years to come, this disease is expected to affect even greater populations, especially as the elderly population grows disproportionately [3]. Glaucoma is known to be multi-factorial in origin, with established genetic and biological risk factors. However, the fundamental causes remain unknown for many types of glaucoma.
Glaucoma is often classified into primary open-angle (POAG), primary angle-closure (PACG), secondary angle-closure, and secondary open-angle, congenital and juvenile glaucomas. The most common type may differ from one region of the world to another. For instance, PACG is more prevalent in certain regions in Asia, whereas POAG is more equally distributed throughout the world and is the most common form of the disease [2]. Vision loss in most forms of glaucoma is related to elevated intraocular pressure (IOP) with subsequent injury to the optic nerve [4].
The secretion of aqueous humor and regulation of its outflow are physiologically important processes for the normal function of the eye. In the healthy eye, flow of aqueous humor against resistance generates an average intraocular pressure of approximately 15mmHg [5]. IOP is necessary to inflate the eye and maintain the proper shape and optical properties of the globe. The basic concept that impairment in aqueous humor outflow results in elevation of the IOP is a central tenet of glaucoma pathology and treatment. Therefore, understanding the complex mechanisms that regulate aqueous humor circulation is essential for improved management of glaucoma.
FUNCTIONAL ANATOMY
The main ocular structures related to aqueous humor dynamics are the ciliary body (the site of aqueous humor production), and the trabecular meshwork and the uveoscleral pathway (the principal locations of aqueous humor outflow).
The ciliary body attaches to the scleral spur and has the shape of a right triangle. Occupying the innermost and anterior most portion of this structure, in a region called pars plicata, are the ciliary processes. The ciliary processes are the sites of aqueous humor production. The ciliary processes been shown to have increased basal and lateral interdigitations, mitochondria and rough endoplasmic reticulum in the non-pigmented ciliary epithelium, a thinner layer of ciliary stroma, and increased numbers of cellular organelles and gap junctions as compared to other regions of the ciliary body [6]. The epithelium of the ciliary processes has two layers: an inner, non-pigmented layer in contact with the aqueous humor in the posterior chamber, and an external, pigmented layer in contact with the ciliary process stroma. The apical surfaces of the two layers lie in apposition to each other [7, 8]. The non-pigmented ciliary epithelium represents the continuation of the retina; the pigmented epithelium, the continuation of the retinal pigmented epithelium [9]. The posterior part of the ciliary body, called the pars plana, has a flatter inner surface and joins the choroid at the ora serrata. Both sympathetic and parasympathetic nerves supply the ciliary body. Parasympathetic fibers come from the Edinger-Westphal nucleus [10] and pterygopalatine ganglion [11]. Sympathetic fibers originate from the cervical superior ganglion and from the carotid plexus [10], and sensory fibers originate from the trigeminal ganglion by way of the ophthalmic nerve.
The limbus is a transitional zone between the cornea and the sclera. On its inner surface is an identation, the scleral sulcus, which has a sharp posterior margin, the scleral spur and an inclined anterior border that extends to the peripheral cornea [12, 13].
The trabecular meshwork is the structure that overpasses the scleral sulcus and converts it into a circular channel, called Schlemm’s canal. The TM is a triangular, porous structure, in cross section, that consists of connective tissue surrounded by endothelium. TM can be divided in three components: uveal meshwork, corneoscleral meshwork and juxtacanalicular meshwork [14]. Sympathetic innervation of the TM originates from the superior sympathetic ganglion. Parasympathetic innervation derives from the ciliary ganglion. Sensory nerves originate from the trigeminal ganglion [15].
The uveal meshwork forms the lateral border of the anterior chamber, extending from the iris root and ciliary body to the peripheral cornea. The uveal meshwork consists of bands of connective tissue, with irregular openings that measure between 25 to 75µm [16].
The corneoscleral meshwork extends from the scleral spur to the anterior wall of the scleral sulcus and is the most extensive portion of the TM. It is composed of perforated sheets that become progressively smaller nearing Schlemm’s canal (Flocks 1956). The corneoscleral meshwork is organized into four concentric layers, viz. from within outwards connective tissue with collagen fiber layer, elastic fiber layer, “glass membrane” layer (delicate filaments embedded in ground substance) and endothelial layer [17-19].
The outermost part of the trabecular meshwork, composed of a layer of connective tissue lined on either side by endothelium, is called the juxtacanalicular meshwork [20]. The central connective tissue layer has variable thickness and is non-fenestrated and the outer endothelial layer comprises the inner wall of Schlemm’s canal [18, 20].
Schlemm’s canal (SC) is comprised of endothelial cells surrounded by connective tissue like a vein. SC possesses internal collector channels and is connected to episcleral and conjunctival veins through the external collector channels, the intrascleral venous plexus, the deep scleral plexus and the aqueous veins [13, 15] (Fig. 1).
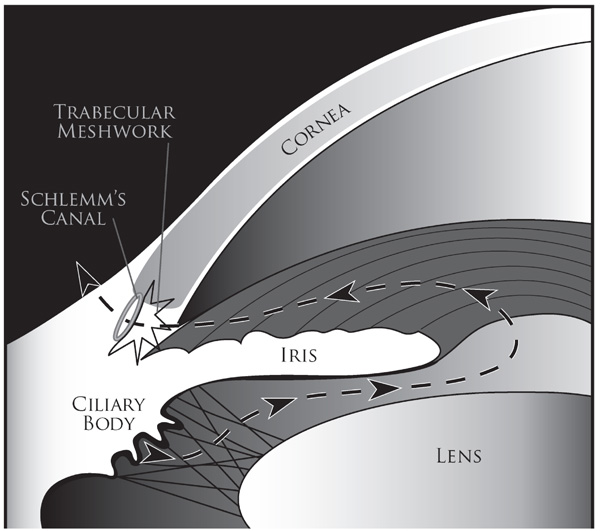
Schematic diagram illustrating the trabecular meshwork conventional outflow pathway. Aqueous humor is produced by the ciliary body and it flows (dashed line shown with arrowheads) from the posterior chamber through the pupil into the anterior chamber. From there it flows out through the trabecular meshwork into the Schlemm’s canal and subsequently absorbed into the episcleral veins via the collector channels.
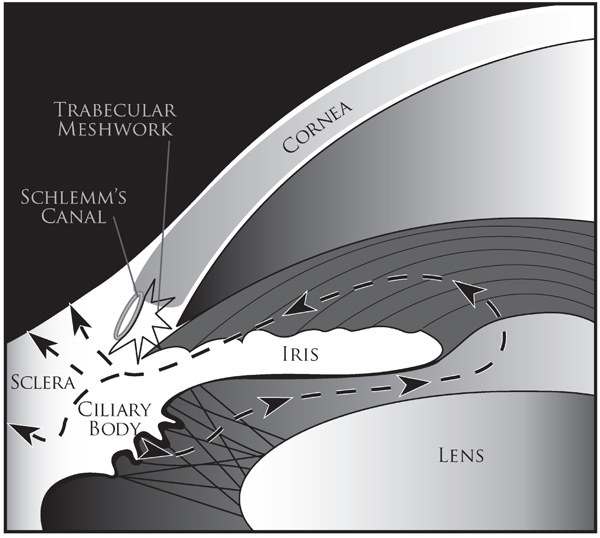
Schematic diagram illustrating the uveoscleral outflow pathway. Aqueous humor is produced by the ciliary body, in uveoscleral route, it flows from the posterior chamber through the pupil into the anterior chamber and then (shown by dashed lines and arrowheads) through the face of the ciliary body and iris root to the ciliary muscle and suprachoroidal space to either veins in the choroid and sclera or through scleral pores to episcleral tissue.
AQUEOUS HUMOR - DEFINITION AND OVERVIEW
Aqueous humor is a clear fluid that fills and helps form the anterior and posterior chambers of the eye. The lens and cornea must remain clear to allow light transmission, and therefore cannot be invested within a vasculature. The aqueous humor is analogous to a blood surrogate for these avascular structures and provides nutrition, removes excretory products from metabolism, transports neurotransmitters, stabilizes the ocular structure and contributes to the regulation of the homeostasis of these ocular tissues. Aqueous humor also permits inflammatory cells and mediators to circulate in the eye in pathological conditions, as well as drugs to be distributed to different ocular structures [21].
Aqueous humor provides a transparent and colorless medium between the cornea and the lens and constitutes an important component of the eye’s optical system. Aqueous humor is secreted by the ciliary epithelium lining the ciliary processes and enters the posterior chamber. Initially, to reach the posterior chamber, the various constituents of aqueous humor must traverse the three tissue components of the ciliary processes - the capillary wall, stroma, and epithelial bilayer. The principal barrier to transport across these tissues is the cell membrane and related junctional complexes of the non-pigmented epithelial layer [22]. Circulating aqueous humor flows around the lens and through the pupil into the anterior chamber. Within the anterior chamber, a temperature gradient creates a convective flow pattern, which is downward close to the cornea where the temperature is cooler, and upward near the lens where the temperature is warmer [23].
The aqueous humor leaves the eye by passive flow via two pathways at the anterior chamber angle, anatomically located at the limbus. The conventional pathway consists of aqueous humor passing through the trabecular meshwork, across the inner wall of Schlemm’s canal, into its lumen, and into draining collector channels, aqueous veins and episcleral veins [24, 25]. The non-conventional route is composed of the uveal meshwork and anterior face of the ciliary muscle. The aqueous humor enters the connective tissue between the muscle bundles, through the suprachoroidal space, and out through the sclera [26, 27].
Equilibrium exists between the production and drainage of aqueous humor. Disruption of aqueous outflow, usually through the conventional pathway, results in elevation of IOP, which is a major risk factor in the pathogenesis of glaucoma [28].
AQUEOUS HUMOR – FORMATION AND COMPOSITION
Three mechanisms are involved in aqueous humor formation: diffusion, ultrafiltration and active secretion [5]. The first two processes are passive and do not entail active cellular participation.
Diffusion occurs when solutes, especially lipid soluble substances, are transported through the lipid portions of the membrane of the tissues between the capillaries and the posterior chamber, proportional to a concentration gradient across the membrane [29].
Ultrafiltration is the flow of water and water-soluble substances, limited by size and charge, across fenestrated ciliary capillary endothelia into the ciliary stroma, in response to an osmotic gradient or hydrostatic pressure [29].
Diffusion and ultrafiltration are responsible for the accumulation of plasma ultrafitrate in the stroma, behind tight junctions of the non-pigmented epithelium, from which the posterior chamber aqueous humor is derived [30, 31].
Active secretion is thought to be the major contributor to aqueous formation, responsible for approximately 80% to 90% of the total aqueous humor formation [32, 33]. The main site for active transport is believed to be the non-pigmented epithelial cells. Active transport takes place through selective trans-cellular movement of anions, cations, and other molecules across a concentration gradient in blood-aqueous barrier. This is mediated by protein transporters distributed in the cellular membrane. Aquaporins (AQPs) are molecular water channels which aid with rapid bulk transport of fluid or transport of fluids against an insufficient osmotic pressure gap. Two AQP’s, AQP1 and AQP4, have been shown to contribute to aqueous humor secretion [34]. The energy required for the transport is generated by hydrolysis of adenosine triphosphate (ATP) to adenosine diphosphate (ADP), which is activated by Na+ and K+ (66) mediated by Na+-K+-ATPase, an enzyme located in both the non-pigmented and pigmented ciliary epithelia [35]. Na+-K+-ATPase can be inhibited by many different molecules, including cardiac glycosides, dinitrophenol [36], vanadate [37], and possibly acetazolamide through pH changes [38]. Thus, Na+-K+-ATPase is of particular interest in pharmacological studies of aqueous humor dynamics.
Another enzyme, carbonic anhydrase, found in the non-pigmented and pigmented ciliary epithelia [39], mediates the transport of bicarbonate across the ciliary epithelium by the reversible hydration of CO2 to form HCO3-and protons through the following reaction: CO2 + H2O ⇌ H2CO3 ⇌ HCO3- + H+ [40]. Bicarbonate formation influences fluid transport by affecting Na+, possibly by regulating the pH for optimal active ion transport [38].
The movement of electrolytes across the ciliary epithelium is regulated by electrochemical gradients and, although there is a net direction of secretion across the epithelium [32], hydrostatic and oncotic forces favor resorption of aqueous humor [41]. Chloride ion is the major anion transported across the epithelium through Cl- channels [35]. Other molecules are also actively transported, including ascorbic acid, which is secreted against a concentration gradient by sodium-dependent vitamin C transporter 2 (SVCT2) [42] and certain amino acids, which are secreted by at least three different solute carriers [43]. Active transport produces an osmotic gradient across the ciliary epithelium, which promotes the movement of other plasma constituents by ultrafiltration and diffusion [44].
The rate of aqueous humor turnover is estimated to be 1.0% to 1.5% of the anterior chamber volume per minute [32], which is 2.4 ± 0.6μl/min (mean ± SD, daytime measurements in adults aged 20–83 years) [45]. Using fluorophotometry, diurnal variations were observed in aqueous humor turnover rates, reflecting a pattern known as the circadian rhythm of aqueous humor flow in humans. Aqueous humor flow is higher in the morning than at night. Aqueous humor flow is normally about 3.0μl/min in the morning, 2.4μl/min in the afternoon, and drops to 1.5μl/min at night [45]. The mechanism that controls this biologic rhythm is poorly understood. Circulating epinephrine available to the ciliary epithelia may be a major driving force [46]. The effect of timolol, epinephrine and acetazolamide on the rate of aqueous humor flow through the anterior chamber has been studied. Epinephrine increased the rate of aqueous flow in sleeping subjects to a greater extent than it did in awake subjects. Timolol reduced the rate in awake individuals, but not in sleeping ones and acetazolamide reduced the rate of flow in both awake and epinephrine-stimulated subjects [47]. Norepinephrine has also been shown to stimulate aqueous flow, but not as effectively as epinephrine [46]. Another hypothesis supporting epinephrine influence on circadian rhythm could be a ciliary production of this hormone. However, epinephrine concentration in human aqueous humor appears to be very low, ranging from 0 to 0.1 ng/ml [48]. Moreover, in patients with surgical adrenalectomy or Horner syndrome (reduced or absent sympathetic innervation on one side), the circadian flow pattern was observed to be normal [49, 50]. Other hormones, such as melatonin, hormones related to pregnancy, and anti-diuretic hormones, do not appear to alter the normal circadian rhythm of the aqueous flow [46]
Measuring the relative concentrations of substances in the aqueous humor and the plasma, as well as in the anterior and posterior chambers, separately make it possible to obtain information about the composition of the aqueous humor [51]. Aqueous humor composition depends not only on the nature of its production, but also on the metabolic interchanges that occur within various tissues throughout its intraocular route [52].
The major components of the aqueous humor are organic and inorganic ions, carbohydrates, glutathione, urea, amino acids and proteins, oxygen, carbon dioxide and water. Aqueous humor is slightly hypertonic to plasma in a number of mammalian species [53-55], except from eyes of rhesus monkeys, in which no significant difference was observed [56]. When comparing anterior chamber and posterior chamber fluids separately, no differences were found in osmolarity, total concentration of dissolved substances or pH [57]. Most studies have shown the Na+ concentration in plasma and aqueous humor to be similar [57, 58]. The greatest differences in aqueous humor relative to plasma, are the concentrations of protein (200 times less) and ascorbate (20 to 50 times higher) [59]. The protein content of aqueous humor has both quantitative and qualitative differences compared to plasma. Most aqueous humor proteins are intrinsic glycoproteins of the vitreous, which are secretory products of the inner epithelial layer of the ciliary body [60]. Specific classes of immunoglobulins, such as IgG, were found to be in a higher concentration in the aqueous humor as compared to IgM and IgA levels [61, 62]. Relative concentrations of free amino acids vary, with ratios to plasma concentration ranging from 0.08 to 3.14, reinforcing the concept of active transport of amino acids [63]. Glucose and urea in the aqueous humor are approximately 80% of the plasma levels. Important anti-oxidant substances can also be found in the aqueous humor, such as glutathione (derived by diffusion from the blood) and ascorbate (which helps protect against light-induced oxidative damage) [32]. A number of molecules involved in the maintenance of the extracellular matrix, such as collagenase, have been identified in human aqueous humor, which may influence TM outflow resistance and, consequently, the IOP [64]. In addition, growth factors have been detected in the aqueous humor, as well as receptors for many of these factors on target tissues, including transferrin [65], transforming growth factors [66], endothelin-1 [67] and indoleamine 2,3-dioxygenase [68].
AQUEOUS HUMOR – OUTFLOW
As mentioned earlier, the aqueous humor exits the eye through both conventional and unconventional pathways. The physiology of these two routes differs in several important ways. For example, the outflow through the unconventional pathway is relatively independent of the intraocular pressure unlike the conventional pathway [69], their behavior in response to different pharmacological agents [70], also rate limiting step in unconventional pathway is flow through the ciliary muscle and in the conventional pathway it’s the flow through the inner wall of schlemm’s canal [71]. Regulation of the extracellular matrix (ECM) composition appears to influence aqueous humor outflow resistance in both the pathways [72].
Another mechanism reported to influence aqueous humor outflow involves age-dependent changes. For instance, in the healthy aging human eye, a reduction in the production of aqueous humor is balanced by a reduction in its drainage through the uveoscleral outflow pathway, thereby leaving intraocular pressure relatively unchanged [73] (Fig. 2).
Fluid movement takes place down a pressure gradient from the TM into Schlemm’s canal and through the inner wall of Schlemm’s canal, following the conventional route, and appears to be a passive pressure-dependent transcellular mechanism, frequently associated with paracellular routes, such as giant vacuoles and pores acting as one-way valves [74]. These pores range in size from 0.1 to 3µm in diameter, and are the main passageway not only for aqueous humor, but also for particulate materials, such as cells, ferritin and microspheres [75-77]. Changes in IOP bring about changes in the structure of the endothelium lining Schlemm’s canal. Elevated IOP leads to an increase in the number and size of these vacuoles, and vice versa [78, 79]. The inner wall of Schlemm’s canal is a complex tissue that is poorly understood - there is still doubt if it influences outflow facility in normal or glaucomatous eyes, even though circumstantial evidence points in that direction [80].
After exiting Schlemm’s canal, the aqueous humor enters the aqueous veins and, subsequently, mixes with blood in the episcleral veins, where the pressure is approximately 8–10 mmHg [81, 82], and the resistance of the conventional aqueous drainage tissues is approximately 3–4 mmHg/µl/min. This results in an average IOP of 15.5 ± 2.6 mmHg (mean ± SD) for the general population [83].
In humans, 75% of the resistance to the aqueous humor outflow is localized to the TM, and 25% occurs beyond Schlemm’s canal [84]. On this basis, trabeculotomy and trabeculectomy were proposed as surgical therapies for treatment of POAG [85]. The major site of resistance within the TM structure has not yet been well characterized, but direct pressure measurements [86, 87] and circumstantial evidence [80] indicate that it resides in the juxtacanalicular portion [88, 89].
Some studies suggest that glycosaminoglycans, which constitute the fundamental substance of the ECM of the TM [90-92], are partly responsible for increased resistance to outflow. The osmotic forces exerted by glycosaminoglycans may induce hydration (edema) of the TM, which can cause obstruction of the trabecular structure [93]. Catabolic enzymes released from lysosomes depolymerize glycosaminoglycans and prevents this obstruction. This effect is also inhibited by corticosteroids, which prevent the release of the enzymes by stabilizing the lysosomal membranes and has been associated with a role in outflow obstruction and glaucoma pathogenesis [93, 94]. In glaucomatous eyes, an increase in the ECM thickness beneath the inner wall of Schlemm’s canal and in the juxtacanalicular meshwork compared with age-matched healthy controls has been observed [95]. Other studies suggest that the interaction of ECM components with different proteins may induce formation of deposits that obstruct aqueous humor outflow through the TM. For instance, proteomic analyses have identified cochlin, a protein of incompletely understood function, in the glaucomatous TM but not in healthy controls. Functionally, cochlin undergoes multimerization induced by shear stress and other changes in the microenvironment. Cochlin, along with mucopolysaccharide deposits, have been found exclusively in glaucomatous TM [96, 97].
The influence of the iris and ciliary muscle, two contractile structures innervated with cholinergic nerves, on the resistance to aqueous outflow has also been cnntemplated. The anterior tendons of the ciliary muscle insert into the outer portion of the corneoscleral meshwork and into the juxtacanalicular meshwork [98]. During contraction, the ciliary muscle moves in an anterior and inward direction, resulting in spreading of the TM and dilation of Schlemm’s canal, thus decreasing outflow resistance. During relaxation, the opposite occurs, thereby increasing outflow resistance [99]. Studies in various animal species demonstrated that voluntary accommodation, electrical stimulation of the trigeminal nerve, and local or systemic administration of cholinergic agents decrease outflow resistance [100-104].
Direct and indirect acting muscarinic cholinergic agonists have been used in the medical management of primary open angle glaucoma. Recent studies demonstrate the presence of at least two different subtypes of muscarinic receptors in the ciliary muscle and in the trabecular meshwork tissue or cell cultures from human eyes [105, 106]. In addition, cholinergic agonists, such as oxotremorine, induce the contraction of the ciliary muscle by binding selectively to receptors located in the longitudinal portion of the muscle, indicating that these agents may modulate the outflow facility independently from accommodation and miosis [105]. Ultrastructural and histochemical differences between the longitudinal (more relevant to outflow facility) and circular (more relevant to accommodation) portions of the ciliary muscle have been observed in monkey eyes. However, these differences in muscarinic receptor subtypes do not appear to play a role in dissociation of accommodative and outflow resistance responses [107, 108].
Uveoscleral outflow was described by observing the exit of radioactive tracers into the anterior chamber of the cynomologous monkey eye [109]. Further characterization of this pathway derives mostly from animal experiments and from mathematical calculations of an expanded Goldmann equation (i.e. F= (Pi-Pe) X C+U where F is the rate of aqueous humor formation, Pi is the intraocular pressure, Pe is the episcleral venous pressure, C is the tonographic facility of outflow and U is the pressure insensitive parameter to symbolize uveoscleral outflow) [109]. This model has been utilized for many years and, in the opinion of many investigators, views the aqueous outflow as passive fluid movement down a pressure gradient [110, 111]. The main obstacles in the assessing more accurately aqueous outflow values with the Goldman equation are: the pressure of the episcleral venous plexus into which aqueous humor flows, and that uveoscleral flow must be independent of the IOP for the equation to be relevant. Although the relationship established is acceptable, it is oversimplified, since this formula implies the recipient pressure for all pressure-dependent outflows can be represented by a single episcleral vessel [69]. In monkeys, uveoscleral flow is not truly pressure-independent and the relationship between flow and IOP is not linear [70]. Ciliary muscle contraction greatly affects uveoscleral outflow [112], and prostaglandin F2α greatly increases uveoscleral outflow by decreasing the flow resistance of the interstitial spaces in the ciliary muscle [113, 114].
Some investigators consider uveoscleral outflow analogous to lymphatic drainage of tissue fluid in other organs, since the fluid may be drawn osmotically into the veins and may mix with tissue fluid from the ciliary muscle, ciliary processes and choroid [27]. In non-human primates, 40-50% of aqueous humor exits the eye by the uveoscleral route. In human eyes, most data has been collected by indirect calculations, with results suggesting a similar fraction, at least in eyes from younger individuals. An age-dependent reduction in uveoscleral flow in human eyes may explain the initial difference seen between non-human primate and human eyes [71]. In eyes treated with atropine, uveoscleral flow accounts for 4 to 27% of the total outflow, but with pilocarpine it was only 0 to 3% [115].
The biomechanics of aqueous humor flow within the anterior chamber also need to be considered. IOP is the loading force to which the outflow system normally responds. Evidence of tissue and cellular deformation in response to an IOP-induced load places TM resistance to IOP at the level of the endothelium of Schlemm’s canal. Schlemm’s canal endothelium attaches tightly to the TM by extending cytoplasmic processes into the juxtacanalicular space [78, 116, 117]. Well-characterized desmosomes, capable of sustaining cellular stress, are present between cell process attachments [118, 119]. The trabecular lamellae contain type I and III collagen, which provide structural support in tension, and elastin, and a recoverable response over large excursions. The organization and distribution of elastin in trabecular lamellae is similar to that found in tendons [120] and enables a mechanism for reversible deformation in response to cyclic hydrodynamic loading [78]. Progressive deformation of Schlem’s canal juxtacanalicular cells and trabecular lamellae with increasing IOP occurs in concert with progressive enlargement of the juxtacanalicular space. This movement causes cellular elements and ECM material to become less compact and progressively reduce the ability of the juxtacanalicular space to participate as a resistance element [78, 119, 121]. However, as IOP increases, resistance to aqueous outflow also increases [122, 123]. This makes the juxtacanalicular region an unlikely source of hydraulic resistance [124]. Trabecular and vascular endothelial cells are mechanosensors [125] that direct vessel wall self-organization [125] in order to optimize wall and shear stress. Pressure and shear stress-mediated signals in endothelia initiate a series of responses at the cellular, molecular, and genetic levels, and induce both rapid responses and slow adaptive changes that regulate pressure and flow [125, 126]. These processes are not linear but are part of a highly complex interactive network [125] in which an alteration in any component requires a contemporaneous adjustment of numerous other components in an interactive fashion resulting in long-term homeostasis [127, 128].
Current pharmacological therapies for lowering the intraocular pressure in glaucoma include increasing aqueous humor outflow and suppression of aqueous humor production. For example, aqueous humor production is reduced by both topical and systemic carbonic anhydrase inhibitors which decrease the production of aqueous humor by the epithelial cells of the ciliary body. Aqueous humor outflow is increased by prostaglandin agonists that increase outflow mainly through the uveoscleral pathway, possibly through the activation of matrix metalloproteinases, and also through the trabecular meshwork. Other glaucoma medications such as adrenergic agonists decrease outflow resistance through mechanisms that are not completely clear.
Current surgical therapies also seek to lower the intraocular pressure by increasing aqueous outflow or decreasing aqueous humor production. For example, ciliary body ablative laser treatments such as cyclcophotocoagulation laser (either transcleral or endopscopic) seek to partially destroy the cililary body to decrease aqueous production. In contrast, laser trabeculoplasty modifies the trabecular meshwork (in a manner still not known) to decrease outflow resistance. Other surgeries such as trabeculectomy, glaucoma drainage implants, and glaucoma shunts (which penetrate the trabecular meshwork and cannulate Schlemm’s canal or create a path through the scleral wall) bypass outflow resistance by shunting aqueous humor through or around the trabecular meshwork. Newer procedures such as canaloplasty (which dilates Schlemm’s canal) and other shunts which seek to open the uveoscleral pathway by creating a mini-cyclodialysis cleft all involve increasing outflow by opening existing aqueous drainage pathways or creating new pathways.
ACKNOWLEDGEMENTS
Some of the pertinent original work in our laboratories were supported by National Institutes of Health Grants R01 EY16112, K08 EY016775, and P30 EY014801; a career development award (SKB) and an unrestricted grant to the University of Miami from Research to Prevent Blindness. We thank Dr. Carol Toris for her helpful comments on a previous version of the manuscript.

