All published articles of this journal are available on ScienceDirect.

The Effect of Changing Colours on Central Crowding Reading
Abstract
Background:
Crowding can be defined as the impaired recognition of closely spaced objects. Changing colour and lighting enhance visual comfort and perceptual troubles that influence impaired vision reading.
Objective:
The current study was aimed to investigate the impact of changing the flanker distance and unflanked targets with colours on central crowding reading for subjects with their distant best correction (BCVA) equal to or greater than 6/6.
Methodology:
Six native English speakers (age: 18–38) who participated in a cross-section intervention study were asked to identify the orientation of the letter E (flanked or unflanked) in different directions around the central target in different colours (red, green, blue and black) on a white background.
Results:
Different colours affect central crowding (p<0.05). However, the central crowding reading of red was not affected by changing flankers (P > 0.05).
Conclusion:
Central reading crowding is visual crowding. Different colours affect central crowding. However, the central crowding reading in red was not affected by changes in flankers.
1. INTRODUCTION
Reading is a complex cognitive process of decoding signs to derive meaning. It is additionally a way of language comprehension, communication, and sharing ideas and information. Hence, reading is an essential necessity for a progressed society. Reading is underlined by three basic processes: printed word recognition, language comprehension, and sight perception [1]. Recognition of the word is considered an essential literacy skill that facilitates access to and processing of written language, as well as affects reading performance. Word recognition can be defined as the ability to automatically and accurately recognise words with and without semantic setting. It is a performance stamp of skilled readers or the ability to accurately recognize printed words [2].
The positional noise and colour affect reading performance, including word recognition and identification for non-amblyopic and amblyopic participants with uncrowded vision [3-6]. Crowding can be described as the inability to discriminate the targets in clutter, and it plays a vital role in developmental changes in reading speed [7]. It can be described by the harmful influence of closing objects on visual discrimination. Additionally, it is described as a curious phenomenon that can be recognized as targets when viewed in isolation and rendered unrecognizable in clutter [8, 9]. In 1963, Takahashi discovered that the visual crowding effect occurs when the target object and flankers are displayed to different eyes. In 1970, Bouma also discovered that the crowding extent is a more or less constant fraction eccentricity of the target object and that crowding happens in a variety of tasks, including letter recognition, vernier acuity, orientation discrimination, stereoacuity, and face recognition. After that, in 1990, Bouma et al. reported that orientation is not influenced by crowding in specific adaptations. Moreover, the impact of crowding on spatial vision possibly occurs in the primary visual cortex [9]. In both foveal and peripheral vision, a letter recognition ability may be degraded by adjacent contours. In the non-central (peripheral) vision, crowding works over a large distance, where the contours of targets and flanking are well separated.
The object is easily recognized when unflanking and becomes unrecognizable if surrounded by other objects. Furthermore, in peripheral (non-foveal) vision, crowding occurs on very large distances (up to 0.5× of the object eccentricity) and for both high and low contrast. In central vision, crowding happens on small distances (4-6 arc/min) or does not occur at all [9, 10]. Moreover, many studies have recently been conducted to raise object recognition processes understanding, which has important clinical implications for subjects with low vision, such as macular degeneration, amblyopia, and dyslexia [9-14]. This study investigated the impact of changing the flanker distance and unflanked targets with variant colours on central crowding reading for subjects who had BCVA of equal or better than 6/6.
2. METHODOLOGY
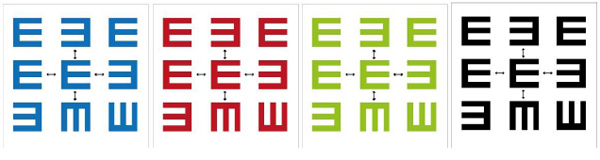
Six participants took part in the cross-section intervention study (4 males and 2 females). They were aged between 18 and 38 years old. Each of the participants had BCVA of equal or better than 6/6. These studies were run on an Apple desktop computer with a MATLAB program, including Psychophysics Toolbox extensions. The experiments were displayed on a Sony screen (21 inches) FD Trinitron with a pixel resolution of 1920 * 1440. The refresh monitor was 75 Hz connected to an Apple desktop computer with an OSX and ATI Randeon HD (5770) 1024 MB graphics card. These experiments were designed in MATLAB, version 2013, 64 bits with a psychtoolbox (3 extensions). A switcher box was used to enhance the luminance depth of the monitor display from 8 bits to 12 bits. General information was taken about the participants, including age, gender, and ocular history. Visual acuity was assessed for each of the participants. In the experiments, the participants responded to the presentation of a stimulus (the letter E) in different directions around the central target. Furthermore, the stimulus was in different colours (red, green, blue, and black) on a white background. The stimulus letter E’s direction was presented randomly and temporarily either singly (unflanked) or surrounded by eight random letters (flanked) with different orientations (0o, 90o, 180o, and 270o) (Fig. 1). The stimulus presentation (E) is threshold acuity in degrees. The threshold is the acuity of E in degrees without any flanking. A computer was used to run the experiment to control the size, spacing, and duration of the stimulus.
In the experiments, the flanker distance was measured at 5 different distances from edge to edge for the target and flanker (Fig. 2). The flanked distances were measured by bar size (stroke width) in minutes of arc.

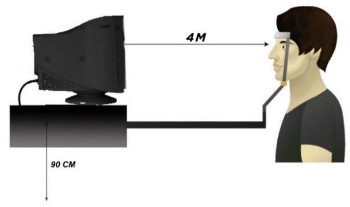
The temporal sequences of the stimulus presentations in the experimental trials were fixed. There was one duration (frame) in all experiments [15]. The refresh monitor was 75 Hz, which means that the duration was 15/75 sec. The experiments were conducted in a dark room. The crowding experiments were run using the staircase method (3 to 1) with the adaptive methods. The participants were asked to fix their vision on the centre of the screen presentation by using keyboard arrows to respond to the target orientation. The distance between the participant and the stimulus presentation was 4 metres (Fig. 3). The stimulus was presented randomly (flanked and unflanked target) with contrast (80%) on a white screen and the participant responded to the stimulus by clicking on the keyboard. The experiment finished when there were either 10 incorrect responses or 60 trials of flanked and unflanked target stimuli. In addition, each stimulus (colour) was examined separately and then followed by other colours for each participant with random colour orders.
3. RESULTS
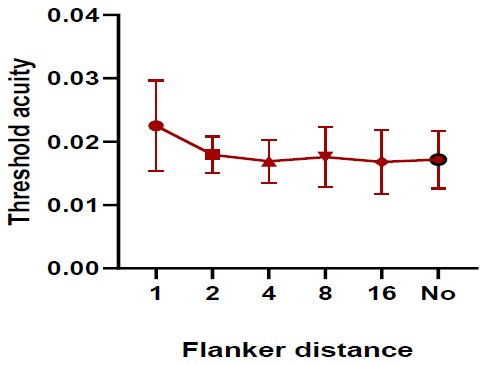
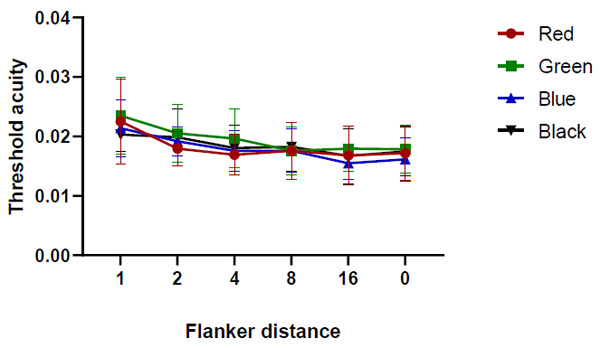
One-way repeated measures ANOVA test indicated that the threshold acuity under the red stimulus was very similar with and without flankers (Pillai’s Trace, F=12.341 and p = 0.213). However, the threshold acuity was insignificantly higher (p > 0.05) when the flanker distance was 1 stroke width (Fig. 4). The results showed that the difference mean was insignificant (one-way repeated measures ANOVA test (Pillai’s Trace, P=0.130, F= 6.96), pairwise comparisons were not statistically significant p > 0.05).
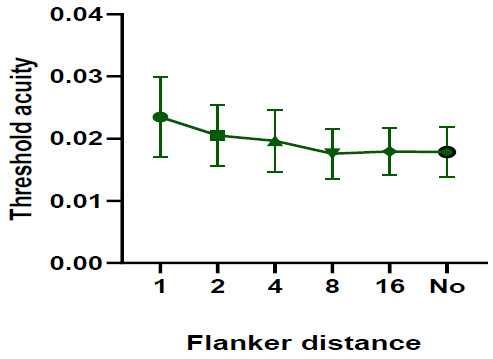
One-way repeated measures ANOVA test (Pillai’s Trace, F=2.635 and p = 0.435) indicated that the threshold acuity under the green stimulus was very similar with and without flankers. However, the threshold acuity was insignificantly higher when the flanker distance was 1 stroke width (Fig. 5). However, the threshold acuity mean difference results were significant between flanker distance 16 and unflanker distance (one-way repeated measures ANOVA test, P=0.048). The results showed that the mean difference in threshold acuity was significant between flanker distance 1 and flanker distance 8 (p = 0.043).
The threshold acuity under the blue stimulus was very similar with and without flankers (one-way repeated measures ANOVA test, F= 69.706 and p = 0.091). However, results showed a significant difference between flanker distance 1 and flanker distance 8, and 16, and flanker distance 8 and flanker distance 16 (p = 0.043, 0.044, and 0.018), respectively. (Fig. 6). The findings indicated that the mean difference in threshold acuity was significant between flanker distance 1 and flanker distances 8, and 16, between flanker distance 2 and flanker distance 16, and flanker distance 4 and flanker distance 16 (p = 0.029, 0.029, 0.049, and 0.012), respectively.
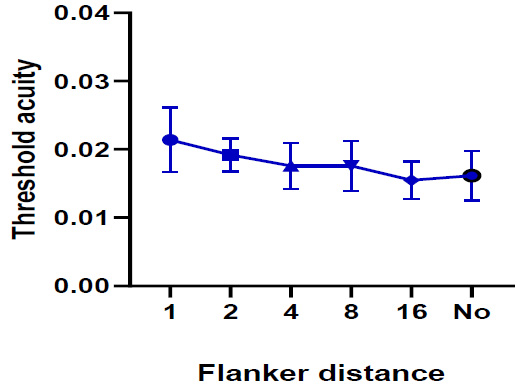
One-way ANOVA repeated measures test (Pillai’s Trace, P=0.081, F= 11.636) indicated that the threshold acuity under the black stimulus was very similar with and without flankers (Fig. 7). However, findings indicated that the mean difference in threshold acuity was significant between flanker distance 2 and flanker distance 16 (p = 0.035).
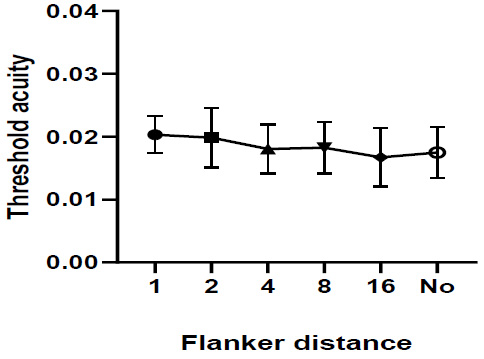
The threshold acuity under all colour stimuli (red, green, blue, and black) was very similar with and without flankers (Two-ways repeated measures ANOVA test (Pillai’s Trace P <0.001, F= 23.036) (Fig. 8). However, multivariate comparisons showed a significant difference between flanker distance 1 and flanker distance 2, 4, 8, 16, unflanker distance (p = 0.024, < 0.001, < 0.001, < 0.001, and < 0.001), respectively, flanker distance 2 and flanker distance 4, 8, 16, and unflanker (p = 0.039, 0.014, < 0.001, and 0.001), respectively.
Two-ways repeated measures ANOVA test (Pillai’s Trace P <0.001, F= 21.363), multivariate comparisons revealed that the mean difference threshold acuity under all colour stimuli (red, green, blue, and black) was very similar with and without flankers (p > 0.05).
4. DISCUSSION
The overwhelming previous studies focused on the crowding peripheral effects and assumed that central crowding was absent [8, 12, 13]. In these studies, the crowding colour reading has shown that there are no obvious differences in the threshold acuities for the four colours (red, green, blue, and black) with and without flankers, which is different from what we expected. These findings were due to the fact that, in foveal vision, each point of the visual field is processed by many overlapping receptive fields. Furthermore, at the fovea centralis of maximum (0.75o), the cortical representation for both V2 and V3 is greater than for V1. As a result, more handling power is devoted to second-level analysis in this area of the visual field. This means that the processing time in V1, when reacting to a small target, can play a part in the crowding process, next to the primary processing in V2 and V3, thereby overcoming the central effect of crowding. Moreover, the central crowding may be overcome by postponed lateral facilitation [13]. However, the threshold acuity at flanker distance 1 was higher than that of other flankers and unflankers. This is presumably because the critical spacing or flankers and the target may physically overlap or attribute to simple masking of lateral contrast instead of the characteristic neural spatial condition that describes crowding [9-11].
The results showed that the mean differences in threshold acuity varied between the colours and flankers. With red, there was no obvious difference in threshold acuity between flanker distances. The findings of the current study are consistent with this Põder (2007), who reported that red letters were more effective in reducing crowding effects than a yellow blob. This is presumably for the role of magnocellular processing in reading performance, which weakens the reading under red longer wavelength light, as claimed by Chase et al. (2003). However, it is hard to explain how this mechanism works with a longer wavelength (red lighting) to reduce the crowding effects.
However, for the other colours (green, blue, and black), there were obvious differences in threshold acuity for the flanker distances. In green, the differences in the threshold acuities between flanker distances were the highest. This is presumably because the threshold acuity reduction appears to be a normal reduction that there is no factor affecting the reduction under the green wavelength light, and the same results were obtained in previous experiments on orthographic readings [14]. In the blue and black colours, the differences in the threshold acuities between flanker distances were obvious; however, the difference in the threshold was less than for green. For blue, this may be attributed to the role of the magnocellular cells and koniocellular cell pathways, improving orthographic reading and word identification by reducing the positional noise effect [15, 16]. For black, it is in the mesopic vision range (a range of human vision with both rods and cones active), which detects any photons of light and can help with word identification and reduce the threshold acuity or noise limitation average difference [17].
The current study revealed the adverse effects of changing colour in central crowding reading with participants who had BCVA 6/6 or better. However, the central crowding reading to red was not affected by changing flankers. There is evidence that changing colour affects central crowding reading.
CONCLUSION
Central crowding reading is visual crowding. Different colours affect central crowding. However, the central crowding reading in red was not affected by changes in flankers.
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
This study was approved by the Ethics Committee of Glasgow Caledonian University.
HUMAN AND ANIMAL RIGHTS
No animals were used in this research. All human research procedures followed were in accordance with the ethical standards of the committee responsible for human experimentation (institutional and national), and with the Helsinki Declaration of 1975, as revised in 2013.
CONSENT FOR PUBLICTION
Informed consent was obtained from all the participants.
AVAILABILITY OF DATA AND MATERIALS
Not applicable.
FUNDING
None.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
ACKNOWLEDGEMENTS
We would like to thank the Optometry Department at Qassim University in Saudi Arabia for the sponsorship of this study. Furthermore, we would like to express our gratitude to the participants who have given their time willingly to participate in our research.

