All published articles of this journal are available on ScienceDirect.

Levocetirizine Inhibits Migration of Immune Cells to Lymph Nodes and Induces Treg Cells in a Murine Type I Allergic Conjunctivitis Model
Authors Info & Affiliations
Abstract
Background & Purpose:
Levocetirizine is a histamine H(1) receptor antagonist. Here, we utilised DO11.10TCR transgenic mice to establish an antigen-specific T cell-dependent allergic conjunctivitis (AC) model to determine the effect of the topical application of an ophthalmic formulation of Levoceritizine as a treatment for AC.
Experimental Approach:
DO11.10 mice (n=6/each) were exposed to ovalbumin (OVA, 50 µg) and treated with a Levocetirizine ophthalmic formulation (0.001–0.02% v/w) or placebo (vehicle) for 24–72 h. Serum, aqueous/vitreous humour and conjunctiva were obtained. Immunoglobulin (Ig)-E, interleukin (IL)-10 and lipoxin (LX)A4 were determined by ELISA. Levels of tumour necrosis factor (TNF)-α, transforming growth factor (TGF)-β, interferon (IFN)-γ and 18rS expression were measured by RT-PCR. Proportions of total and activated antigen-presenting cells (APC), recruited T lymphocytes (CD4+), activated T lymphocytes (CD25+) and T regulatory cells (Treg) were measured by flow cytometry.
Key Results:
OVA exposure induced AC in the animal model indicated by increased expression of LXA4, TNF-α and TGF-β. Levocetirizine treatment (0.01–0.02% v/w) reduced LXA4 in the eye humours. This treatment approach increased systemic IL-10 secretion and reduced TNF-α and TGF-β expression in conjunctiva without changing IFN-γ expression. Levocetirizine reduced APC levels in draining lymph nodes but increased the proportion of total lymphocytes recruited and their differentiation to Treg cells.
Conclusions & Implications:
Levocetirizine effectively reduces the activation and migration of APC to local draining lymph nodes and induces differentiation of Treg cells as one possible mechanism of its anti-inflammatory action.
INTRODUCTION
Allergic Conjunctivitis
Allergic conjunctivitis (AC) is the most common ocular allergy and varies from mild types of AC, such as seasonal and perennial types, to more severe diseases such as atopic keratoconjunctivitis and vernal keratoconjunctivitis, leading to impaired vision [1].
AC is initiated by the activation and polarisation of the immune response to environmental agents, culminating in the generation of a Th2-dominant immune response and the preferential generation of IgE antibodies. The early phase response occurs when allergens engage specific IgE antibodies bound to FcεRI receptors on conjunctival mast cells. Cross-linking of IgE antibodies triggers the degranulation of mast cells and the release of histamine. EPR is followed by a late phase reaction (LPR), 6–12 h later, and involves the infiltration of inflammatory cells, especially eosinophils [2,3]. Antigen-specific T cells initiate eosinophil infiltration into the conjunctiva, which can result in tissue damage [4].
Dual-action ophthalmic medications (antihistamine and mast cell stabilisers) are associated with improved adverse event profiles but also with side effects of increased duration. These side effects include eye irritation, conjunctival cysts, foreign body sensations, bitter taste, anxiety, local toxicity, secondary open-angle glaucoma, cataracts, and superinfections [1,5,6].
Levocetirizine is a late-generation selective antihistaminic, that acts through H1 receptor. The medication is licensed for the symptomatic treatment of seasonal allergic rhinitis, perennial/persistent allergic rhinitis and chronic idiopathic urticaria [7]. Levocetirizine exhibits antihistaminic effects characterised by rapid onset and extended duration. The drug is also characterised by rapid absorption and high bioavailability, a low potential for drug interactions, low-volume distribution and a lack of effects on cognition, psychomotor function or the cardiovascular system [8]. Levocetirizine inhibits eotaxin-induced eosinophil transendothelial migration through monolayers of human dermal or lung microvascular endothelial cells in vitro. The drug also inhibits both resting and granulocyte-macrophage colony-stimulating factor (GM-CSF)-stimulated eosinophil adhesion to vascular cell adhesion molecule-1 (VCAM-1), eotaxin production by endothelial cells and ICAM-1, as well as major histocompatability complex (MHC) class I expression by interferon (IFN)-γ-stimulated keratinocytes [9]. In addition, the drug modulates the histamine-dependent release of GM-CSF and chemokines. Levocetirizine reduces interleukin (IL)-7 and stem cell factor release by lipopoylsaccharide-stimulated eosinophils [9]. Physiologically-relevant concentrations of Levocetirizine inhibit ICAM-1 expression and secretion of IL-6 and IL-8, as well as nuclear factor (NF)ĸ-B activation [10]. More recently, Levocetirizine was evaluated for its ability to inhibit allergen-induced wheal and flare reaction from 1.5 to 24 h after administration of the drug [11].
Based on the extensive history of clinical effectiveness and tolerability of oral Levocetirizine in the treatment of allergic symptoms, Levocetirizine is being developed as an ophthalmic medication for allergic conjunctivitis. In order to test the anti-allergic activity of ophthalmic formulations and determine their possible mechanisms of action, several animal models have been developed to improve the analysis of molecular and cellular events. Thus, Shii, et al. [12], established a transgenic system in which virtually all thymocytes and peripheral T cells express the transgenic T cell receptor (TCR) from a T cell hybridoma, DO11.10, which recognises chicken ovalbumin (OVA) in the context of I-Ad. This model has been used to analyse blepharoconjunctivitis, uveitis, and type 1 allergic conjunctivitis [12]. In this report, the authors state the benefits of studying the response of the conjunctiva to an immune challenge recreating the allergic response.
In this study, we utilise DO11.10 TRC transgenic mice to establish an antigen-specific T cell-dependent AC model, as previously described [12-14], to determine the effect of topical application of an ophthalmic formulation of Levoceritizine as a treatment for allergic conjunctivitis and the effect of this treatment on the inflammatory response at a cellular level.
METHODS
Mice
DO11.10 TCR transgenic mice (balb/c background, Jackson Laboratories, Bar Harbor, ME), between 6 and 8 weeks of age, were maintained in a pathogen-free environment at the FES-Iztacala, U.N.A.M. animal facility. This, in accordance with the Guide for the Care and Use of Laboratory Animals, as adopted and promulgated by the U. S. National Institutes of Health and the Mexican Regulation of Animal Care and Maintenance (NOM-062-ZOO-1999, 2001). We also followed the guideline in the Statement for the Use of Animals in Ophthalmic and Visual Research as published by the Association for Research in Vision and Ophthalmology.
OVA-Induced Ocular Allergy and Treatments
Ocular allergy in mice was induced using a topical solution of OVA (50 µg for each eye), and sodium fluorescein (FITC, 1 µg in PBS for each eye) was used as a cell marker of uptake and activation of antigen presenting cells (APC). Six h after OVA exposure, eyes were treated every 6 h for a duration of 24 h and then every 12 h for 2 more days with 10 µl per eye of an ophthalmic formulation of Levocetirizine (0.001, 0.005, 0.0, 0.015 and 0.020% v/w) or vehicle (placebo; the same vehicle used in the ophthalmic formulation but without the Levocetirizine). Endpoints were evaluated 24 and 72 h after treatment (n=6). Mice were sacrificed by CO2 inhalation; serum was obtained from cardiac puncture; vitreous and aqueous humour and conjunctival samples were collected from each eye and local draining lymph nodes were collected.
Determination of IgE, IL-10 and Lipoxin A4 (LXA4)
Serum was obtained from blood and quantitative analysis of IgE and IL-10 levels were determined using the BD OptEIATM Set (BD Biosciences Pharmingen, San Diego, CA, USA), according to the manufacturer’s recommendations. Briefly, samples (25 µl) were diluted with PBS (75 µl) and added to the microplate previously coated with capture antibody. Detection Ab + SAv-HRP were incubated at room temperature (RT) for 1 h. The bound enzyme conjugate was detected by adding substrate solution. Absorbance was detected with a microplate reader at 650 nm [15].
Quantitative analysis of LXA4 levels in vitreous and aqueous humour was performed by ELISA (EA 45 Lipoxin A4 assay kit, Oxford Biomedical Research), following the manufacturer’s instructions. Briefly, samples (5 µl) were diluted with extraction buffer (20 µl) and added to microplates. The enzyme conjugate was incubated at RT for 1 h and detected by adding substrate. Absorbance was detected at 650 nm.
Quantification of mRNA by Semiquantitative RT-PCR
Total RNA was extracted from the tissue of each mice using Trizol reagent, according to the manufacturer’s recommendations. Specific mRNA levels were determined by semiquantitative RT-PCR using 18S rRNA as an internal control (all chemicals were Invitrogen). Briefly, 2 µg of RNA were incubated with 250 ng of random primers and 200 U of reverse transcriptase (SuperScript III) for 30 min at 50º C. The PCR was conducted under the conditions indicated in Table 1 with denaturation for 50 s at 94º C, primer annealing for 40 s, extension for 40 s at 72º C, and 7 min extension at 72º C. Ten microlitres of the PCR products were separated by electrophoresis on a 2% agarose gel. Gels were stained with ethidium bromide, and PCR products were quantified by densitometry with Image J software (http://rsb.info.nih.gov/ij/). RT-PCR products were normalised per amount of 18rS product [16].
Primer Sequences, Size Products, and Conditions Used in RT-PCR Assays
| Target | Primer (5´- 3´) | Size (bp) | Annealing Temp/Cycles (ºC) | Accession Number |
|---|---|---|---|---|
|
|
||||
| 18rS | 1F-GGACAGGATTGACAGATTGATAG | 101 | 60/12 | R003278.1 |
| R-CTCGTTCGTTATCGGAATTAAC | ||||
|
|
||||
| TNF-α | F-GGCAGGTCTACTTTGGAGTCATTGC | 307 | 59/40 | NM013693.2 |
| R-ACATTCGAGGCTCCAGTGAATTCGG | ||||
|
|
||||
| TGF-β | F-GCCCTTCCTGCTCCTCAT | 206 | 52/40 | NM011577.1 |
| R-TTGGCATGGTAGCCCTTG | ||||
|
|
||||
| IFN-γ | F-AGCGGCTGACTGAACTCAGATTGTAG | 247 | 52/45 | NM008337.3 |
| R-GTCACAGTTTTCAGCTGTATAGGG | ||||
1 F; Forward
R; Reverse.
Flow Cytometry Analysis
In order to determine drug treatment efficacy, single cell suspension prepared from draining lymph nodes were analysed by fluorescence activated cell sorting (FACS). Monocytic MHC-II expression was determined using monoclonal antibodies PE-Rat anti-mouse 1-A/1-E (BD Biosciences Pharmingen, San Diego, CA, USA) and CD4+CD8- TCRhI cells were detected using PerCP anti-mouse TCR DO11.10 antibody (BioLegend San Diego, CA, USA). Briefly, 1×106 cells were incubated with 100 µl of 1:200 diluted antibodies for 30 min in the dark. Cells were washed in PBS and analysed with a FACS-Calibur flow cytometer, using Summit 3 [17,18].
We utilised a Mouse Treg Flow kit (Biolegend, 320018) to stain Treg cells following the manufacturer’s instructions. Briefly, 106 lymph node cells were incubated for 30 min at 4° C with rat anti-mouse antibodies against FOXP3-Alexa Flour 488/CD4-APC/CD25-PE or rat anti-mouse IgG1κ as a control isotype. After staining, cells were washed twice with PBS with 5% FBS and 0.02% sodium azide and 20000 events per sample were evaluated by flow cytometry.
Statistical Analyses
Data are presented as mean ± standard deviation (SD). Statistical significance was assessed using the Student´s t test; differences were considered significant at p≤0.05 *vs positive control (OVA treatment alone), avs placebo, bANOVA (post-hoc Bonferroni).
RESULTS
Parameters of Inflammation in Serum and Aqueous/ Vitreous Humours
OVA exposure in this animal model did not modify IgE levels after 24 h, but significantly reduced IgE levels after 72 h. Placebo reduced IgE levels only after 24 h (Fig. 1A). Levocetirizine treatment increased IgE levels after 24 h but showed almost no effect after 72 h of treatment when compared to the placebo. Exposure to OVA or placebo did not significantly modify IL-10 levels, except for the higher dose, which increased IL-10 secretion (Fig. 1B). We observed an increase of LXA4 in the aqueous/vitreous humours when animals were exposed to OVA (Fig. 1C). Treatment with placebo reduced LXA4 significantly after 72 h, and treatment with Levocetirizine showed a dual effect. Compared with placebo treatment, Levocetirizine increased LXA4 levels at concentrations of 0.001–0.01% but reduced LXA4 levels at the highest doses (0.015 and 0.02%) after 24 and 72 h, thereby exhibiting a dose-dependent effect at the highest concentrations used.
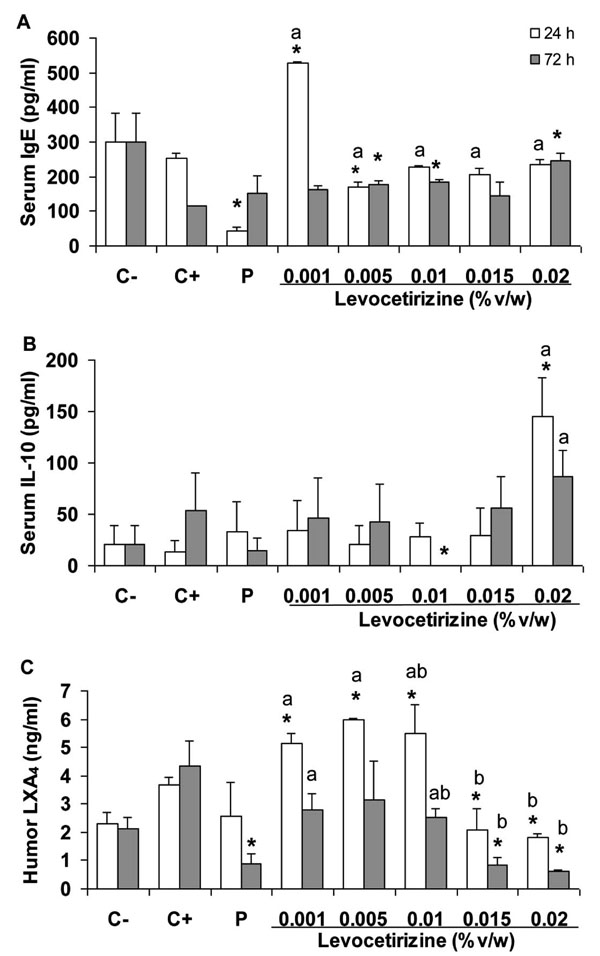
IgE (A) and IL-10 (B) levels in serum, and LXA4 levels in aqueous/vitreous humour (C) (ELISA) from DO11.10TcR-Tg mice (C-) exposed to OVA for 6 h (C+) and treated for 24 (white bars) or 72 h (grey bars) with placebo (P) or Levocetirizine ophthalmic solution at five different concentrations. *p<0.05 vs C+, ap<0.05 vs placebo, Student’s t-test (n = 6), bp<0.05 ANOVA.
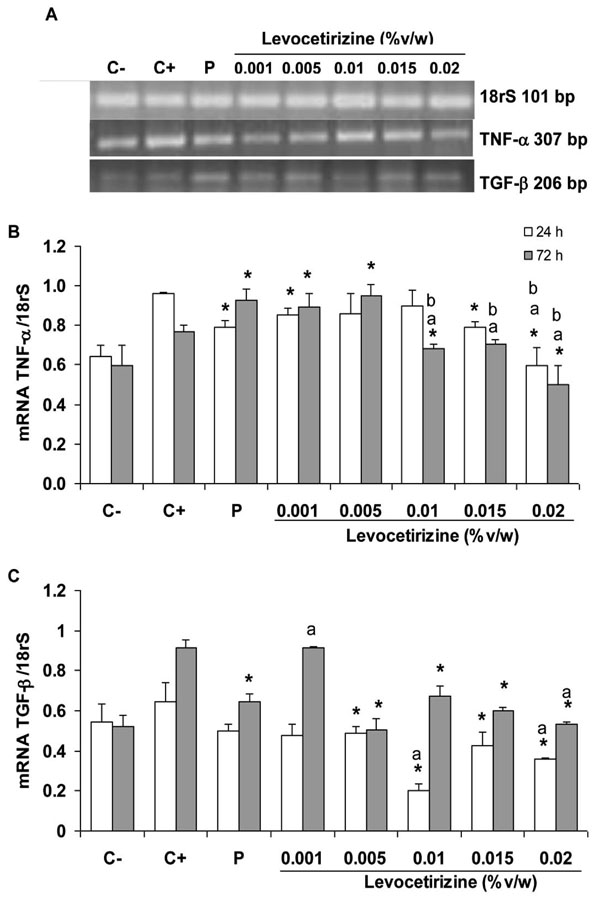
Relative mRNA expression (A) of TNF-α (B) and TGF-β (C) levels in conjunctiva from DO11.10TcR-Tg mice (C-) exposed to OVA for 6 h (C+) and treated for 24 (white bars) or 72 h (grey bars) with placebo (P) or Levocetirizine ophthalmic solution at five different concentrations. mRNA of 18rS was utilised as an internal control of gene expression. *p<0.05 vs C+, ap<0.05 vs placebo, Student’s t-test (n = 6), bp<0.05 ANOVA.

Proportion of total monocytes (A), OVA-activated APCs (B), and recruited T lymphocytes (C) in draining lymph nodes (flow cytometry) from DO11.10TcR-Tg mice (C-) exposed to OVA for 6 h (C+) and treated for 24 (white bars) or 72 h (grey bars) with placebo (P) or Levocetirizine ophthalmic solution at five different concentrations. *p<0.05 vs C+, ap<0.05 vs placebo, Student’s t-test (n = 6), bp<0.05 ANOVA.

Induction of Treg cells in the draining lymph nodes from DO11.10TcR-Tg mice (C-) exposed to OVA for 6 h (C+) and treated for 72 h with placebo (P) or 0.02% Levocetirizine ophthalmic solution for 72 h, as determined by flow cytometry of lymphocytes in R1 as in the panel to the left. From these cells we gated the CD4+/CD25+ cells (R2), and analysed the Foxp3+cells from these last gated cells (R3). *p<0.05 vs C+, ap<0.05 vs placebo, Student’s t-test (n = 6).
Alteration of Gene Expression of Inflammatory Proteins
TNF-α expression in the conjunctiva was induced by OVA exposure (Fig. 2A). Placebo initially reduced TNF-α expression after 24 h, but increased this parameter after 72 h (Fig. 2B). Treatment with low doses of Levocetirizine (0.001–0.005%) did not modify the effect observed with placebo, but higher doses of Levocetirizine (0.1–0.2%) reduced the expression of TNF-α in a dose-dependent manner (bp<0.05, ANOVA). Changes in the expression of TGF-β in the conjunctiva were not dose-related (bp>0.05, ANOVA, Fig. 2C). OVA exposure increased TGF-β levels; placebo treatment moderately decreased TGF-β levels. Levocetirizine (0.005–0.02%) reduced TGF-β expression to levels similar to those achieved by treatment with a placebo. Only 0.02% Levocetirizine significantly decreased TGF-β levels compared to placebo. As expected in this Th2-differentiated environment, IFN-γ was not produced in significant amounts by the conjunctival cells. Therefore IFN-γ was not statistically modulated by the different treatments (data not shown).
Activation of Antigen Presenting Cells
Due to several reports indicating that IL-10 and TGF-β usually suppress CD4+ T cell activation, inhibiting the inflammatory response, we evaluated the activation and migration of APC to draining lymph nodes, as well as the recruitment of T CD4+ lymphocytes to local draining lymph nodes. The number of total cells recovered from the draining lymph nodes did not differ between treatments (Fig. 3A). OVA exposure increased the total number of APCs in the draining lymph nodes; placebo did not modify the APC migration induced by OVA (Fig. 3B). Levocetirizine treatment did not show a dose-related effect on the total number of APC in the draining lymph nodes (bp>0.05, ANOVA), but most doses of Levocetirizine reduced the total amount of APC in the draining lymph nodes after 72 h of treatment. OVA exposure dramatically increased the proportion of activated APC (relative to the total number of APC) in the draining lymph nodes after 72 h (Fig. 3B). Placebo treatment reduced the proportion of activated APC in the draining lymph nodes at 24 and 72 h. Levocetirizine reduced the proportion of activated APC in the draining lymph nodes, but only at the highest concentrations tested (bp>0.05, ANOVA).
Recruitment of T Cells to the Lymph Nodes
A complementary parameter to the extent of APC activation is the number of recruited lymphocytes in situ, as lymphocytes are the final effector cells. OVA exposure increased the total number of locally recruited T cells in the draining lymph node and placebo did not modify this response (Fig. 3C). Only the higher concentrations of Levocetirizine increased the total number of T lymphocytes in the draining lymph nodes. Therefore no dose-related effect was evident with Levocetirizine treatment (bp>0.05, ANOVA).
Treg Cells Differentiation
The reduction on the proportion of activated APC together with the increase in the total number of locally recruited lymphocytes in the draining lymph nodes suggested the possibility that activated T cells could be differentiating into Treg cells. We determined the proportion of CD4+/CD25+/Foxp3-positive cells (Treg cells, Fig. 4, R3) from that of the total CD4+/CD25+ positive cells in the draining lymph nodes by gating CD4+ cells and then CD25+ cells (R2). We observed that Treg cells were increased by OVA exposure. Placebo treatment slightly reduced the OVA-mediated induction of Treg cells (Fig. 4). Treatment with Levocetirizine 0.02% for 72 h reduced the proportion of CD4+/CD25+ cells, as previously described [19] (data not shown) and increased the differentiation of CD4+/CD25+ cells to Treg cells.
DISCUSSION
We used the DO11.10 animal as a model of allergic conjunctivitis by topically challenging the eyes with OVA. This particular transgenic model responds constitutively to OVA challenge [13,14], thus imitating an allergic response with no need of previous immunisation and sensitisation. The allergic response we observe in the eye is representative of systemic responses in the animal model, particularly to that occurring in the draining lymph nodes. Regulation of T cell-dependent immune responses (such as allergic responses), is mediated in part by APC that process and present antigens to T cells, and secrete cytokines and other chemokines. The anterior chamber of the eye is lined by the corneal endothelium (which rests on a stroma and epithelium that is devoid of class II MHC + APC) and iris/ciliary body (which contain significant numbers APCs) [20]. As such, once the antigen responsible of the allergic reaction (OVA, in this case) has been taken by APCs in the tissues, these APCs migrate to local lymph nodes to present the processed antigen to reactive T cells. They, in turn, activate themselves and secrete cytokines and other chemical mediators that are able to induce inflammatory reactions in the site of the antigenic challenge. All these movements and responses are mediated by cytokines and chemokines that travel trough the blood stream, and thus, modifying serum levels of these chemical mediators. Although the clinical signs of an immune response are mostly local, the immune response is generally systemic, making possible to identify the presence of chemical mediators and activated immune cells in the blood stream.
As previously reported in the DO11.10 mice, we confirmed that this model is not responsive to IgE changes [21] and that there is no production of IFN-γ in the conjunctival cells of this animal model of allergic reactions [22]. Nevertheless, the rest of the parameters evaluated here presented changes in response to OVA exposure that were similar to those observed in other animal models of allergic conjunctivitis. In some cases, when the LPR is IgE-independent, this response is usually dependent on TNF-α, inducing a strong Th2-type response [23]. Our data indicate that this might be the case, as the ocular allergic reaction induced by OVA proved to be IgE-independent and we observed an increase in TNF-α expression. Although it is generally accepted that IL-10 inhibits mast cell degranulation [24] and that IL-10 together with high levels of TGF-β inhibits inflammatory responses by suppressing CD4+ T cells [25], these cytokines do not have immunosuppressive roles in some models of experimentally induced allergic conjunctivitis [26]. Moreover, these cytokines are implicated in wound healing and the maintenance of homeostasis in the eye [27].
In our study, increased IL-10 production occurred only with the highest dose of Levocetirizine used and was not accompanied by an obvious increase in TGF-β expression. On the contrary, a decrease in TGF-β expression was associated with a high level of IL-10 secretion as well as a reduction in TNF-α expression, indicating that Levocetirizine presents a very specific mechanism of action, as it does not reduce all the parameters of inflammation but rather just those related to the induction of Treg cells. These three particular cytokines are involved in the generation of Treg cells when TGF-β production induces IL-10 and inhibits TNF-α. This particular cytokine profile, together with low levels of IFN-γ, induces Treg cells [28], as observed in our study. In fact, TGF-β and IL-10 have been shown to be the immunosuppressive cytokines in immune tolerance models. Specifically, TGF-β1 is implicated in the regulation of autoimmune and inflammatory diseases through the conversion of CD4+/CD25+ T cells into Treg cells [27]. Recent studies in mouse models of allergic conjunctivitis suggest that Treg cells also influence the LPR of allergic conjunctivitis [3].
Levocetirizine increased the proportion of lymphocytes present in the draining lymph nodes, reduced the proportion of CD4+/CD25+ cells, and induced the differentiation of CD4+/CD25+ cells to Treg cells. These results indicate that most of the lymphocytes present in the draining lymph node are probably anergic. An observation that supports this hypothesis is that Levocetirizine also dramatically reduced the proportion of activated APC that migrated from the conjunctiva to the draining lymph node. It is recognised that histamine influences the profile of proinflammatory and immunoregulatory cytokines produced by blood monocytes, tissue macrophages and dendritic cells. This sequence of events polarises immature dendritic cells, transforming them to Th2 response-promoting cells [29]. It has been demonstrated that differential patterns of histamine receptor expression on Th1 and Th2 cells determine reciprocal T-cell responses following histamine stimulation. Th1 cells show predominant expression of H1 receptors, while Th2 cells show increased expression of H1 receptors. Most allergic reactions develop a decrease in the H1:H2 receptor ratio in T cells, and Levocetirizine has proven to prevent this decrease, thus facilitating control of the inflammatory response by targeting H1 receptors [28].
Finally, we observed important changes in the production of LXA4 in the eye chamber when Levocetirizine was applied. The eye actively maintains an anti-inflammatory and immunosuppressive microenvironment via soluble factors such as LXA4. Although LXA4 has, in general, an anti-inflammatory function, in this model of allergic conjunctivitis, we observed a decrease in LXA4 associated with decreased expression of the rest of the inflammatory parameters evaluated. Most probably, this anti-inflammatory protein is playing a different role in the animal model evaluated, as LXA4 has been actively implicated in wound healing rather than in the inhibition of leukocytes [30]. There are no reports indicating the specific role of LXA4 in regulation of the inflammatory process in the vitreum/aqueous humours in the eye or on the effect of Levocetirizine on the regulation of 12/15-LOX. Thus, the relevance of this particular finding deserves further evaluation in the future.
Levocetirizine presents unique mechanisms of action, as it induces the differentiation of Treg cells and Th2-prone APCs in the lymph nodes. These effects limit the local inflammatory response via LXA4. Low doses of Levocetirizine do not exhibit a clear effect on most evaluated parameters of inflammation, but high doses of Levocetirizine (particularly, 0.02% v/w Levocetirizine) reduced all inflammatory parameters and increased the anti-inflammatory parameters evaluated in this study. All these results indicate that topically applied Levocetirizine in an ophthalmic formulation could effectively reduce allergic conjunctivitis; thus, topically applied Levocetirizine represents a good therapeutic option in the arsenal of antiallergic treatments.
CONFLICT OF INTEREST
Authors Garcia-Zepeda SP, Estrada-Muniz E, Elizondo G, Terrazas LI, Rodriguez-Sosa M, Quintana-Hau JD, and Vega L., have no conflict of interest regarding this manuscript. Authors Tornero-Montano R, and Baiza-Duran L., were employees of Laboratorios SOPHIA.
ACKNOWLEDGEMENTS
This work was funded by Laboratorios SOPHIA SA de CV, Mexico.

